
Луиз Фернандо Бертолучи, MD, Бразильская ассоциация Рольфинга, Сан-Паулу, Бразилия
Bertolucci, Luiz Fernando. (2011). Pandiculation: Nature’s way of maintaining the functional integrity of the myofascial system?. Journal of bodywork and movement therapies. 15. 268-80. 10.1016/j.jbmt.2010.12.006.
Аннотация
Потягивание — это непроизвольное растяжение мягких тканей, которое происходит у большинства видов животных и связано с переходами между биологическими поведенческими циклами, особенно с ритмом сна и бодрствования (Walusinski, 2006). Зевание является частным случаем потягивания, воздействующим на мышцы полости рта, дыхательной системы и верхнего отдела позвоночника (Baenninger, 1997). Комбинированное поведение, при котором, как это часто случается, зевание происходит одновременно с потягиванием других частей тела (Bertolini and Gessa, 1981, Lehmann, 1979 and Urba-Holmgren et al., 1977), называют синдромом потягивания-зевания (СПЗ).
СПЗ связан с функцией активации ЦНС, который, как полагают, переводит центральную нервную систему в состояние пробуждения после периода сна и готовит животное к ответу на стимулы окружающей среды (Walusinski, 2006).
В данной статье изучается гипотеза о том, что СПЗ может также участвовать в саморегуляции опорно-двигательной системы, поддерживая способность животного воспроизводить скоординированное и комплексное движение за счет регулярного восстановления и возврата в исходное положение структурного и функционального равновесия миофасциальной системы. В настоящее время признано, что миофасциальная система является интегративной, соединяющей все части тела, поскольку сила мышцы передается через фасциальные структуры далеко за пределы сухожильного крепления самой мышцы (Huijing and Jaspers, 2005). В статье утверждается, что потягивание может сохранить интегративную роль миофасциальной системы за счет a) развития и поддержания надлежащих физиологических связей между фасциальными соединениями и б) изменения предстрессового состояния миофасциальной системы путем регулярной активизации тонической мускулатуры.
Представленные здесь идеи изначально возникли из клинических наблюдений в ходе практики мануальной терапии, которая называется репозиционированием мышц (РМ) (Bertolucci, 2008, Bertolucci and Kozasa, 2010a and Bertolucci, 2010b). Эти наблюдения были дополнены обзором литературы по данному вопросу.
Предложена возможная связь между СПЗ и РМ: Нервные рефлексы, которые обычно возбуждаются при РМ, напоминают рефлексы СПЗ. Это позволяет предположить, что РМ может стимулировать частичные реакции СПЗ, и указывает на один из возможных механизмов действия РМ.
Ключевые слова: фасции; соединительная ткань; миофасции; зевота; ритм сна и бодрствования.
Примечание по терминологии
Термин потягивание (англ. pandiculation от лат. pandiculare — растяжение) чаще всего используется для того, чтобы обозначить как зевание, так и непроизвольное потягивание соматических мышц, которые в совокупности образуют синдром потягивания-зевания (СПЗ). Хотя зевание и потягивание являются связанными процессы, каждый из них может происходить независимо от другого. Большинство исследований СПЗ фокусируются на зевании, хотя иногда также описывается сопровождающее его потягивание. Намного реже встречаются исследования, где внимание фокусируется только на компоненте потягивания. В данном тексте СПЗ будет использоваться для обозначения комбинированного поведения, а термин «потягивание» будет использоваться для обозначения только растягивания. Хотя такое использование не распространено, автор отказался от термина «растягивание» для обозначения непроизвольного растягивания как части СПЗ, потому что его можно спутать с произвольным удлинением мышц, которое происходит при занятиях разными видами спорта и при работе с телом, и представляет собой совершенно отдельное явление.
Потягивание: формирование и поддержание нейронной сети и периферических моторных эффекторов
Потягивание является древним и практически повсеместно распространенным типом поведения, которое отмечается в схожих формах и условиях у многих видов животных (Baenninger, 1997). Регулярность и сила движений при потягивании заставляет предположить, что они могут быть физиологически значимыми. Валюсински отмечает, что, согласно концепциям Дарвина, поведение с высокими метаболическими затратами должно компенсироваться каким-то адаптивным преимуществом (Walusinski, 2006).
Действительно, филогенез и онтогенез потягивания демонстрируют свою вероятную роль в развитии и поддержании моторных функций как в структурных, так и в нервных аспектах. Фрейзер (Fraser, 1989a), основываясь на ультразвуковых исследованиях зародышей овец, упоминает о потягивании плода как механизме, который влияет на функциональное определение движущихся частей костно-мышечной системы и способствует развитию и обслуживанию суставных сочленений. Он также выявил и такую функцию потягивания как самоподдержание тела, которая восстанавливает мышечный гомеостаз у домашних птиц, собак, кошек, лошадей, и у других животных (Fraser, 1989a). У страусов аналогичное «поддерживающее» потягивание описали Сауэр и Сауэр (Sauer and Sauer, 1967).
У человека потягивание также отмечается еще до рождения, начиная уже с 12-й недели созревания плода (de Vries et al., 1982). Его связывают с развитием нейронных сетей (Lagercrantz and Ringstedt, 2001, Marder and Rehm, 2005 and Briscoe and Wilkinson, 2004) и сопутствующих костно-мышечных эффекторов (de Vries et al., 1982 and Walusinski et al., 2005). В действительности, отдельные мышцы начинают отличаться от примитивных мезодермальных конденсаций только после создания нейронных связей (Sadler, 1995), основанных на их конкретных двигательных актах, как это будет описано далее.
Развитие мышц начинается с образования примитивных миобластов, которые соединяются и дают начало первичным мышечным трубкам. Примерно на 10-й неделе беременности в первичные мышечные трубки прорастают двигательные нейроны спинного мозга, которые наделяют их сократительной способностью. Внутри первичных мышечных трубок объединяются дополнительные миобласты, которые в конечном счете порождают вторичные мышечные трубки, на этом этапе они используют общую базальную мембрану. Позднее, в процессе развития плода, вторичные мышечные трубки соединяются с сухожильными структурами. На этом этапе они развивают собственные базальные мембраны и становятся независимыми структурами (Jones et al., 2004).
Дифференциация первичных и вторичных мышечных трубок на популяции раздельных двигательных единиц происходит за счет дифференциации покровных слоев фасций и поверхностей скольжения, которые ограничивают свое содержимое и обеспечивают его независимое движение. Так, на базе общей функциональной основы, формируются дискретные фасциальные выстилки и сетевидные матрицы между ними, позволяющие им скользить относительно друг друга.
Такое механически-структурное сопряжение не ограничивается мягкими тканями. Оно может доходить до кости (Wolff, 1986), до форм суставной поверхности (Kapandji, 1987) и практически до каждой ткани, которая испытывает механическую нагрузку (Moore, 2003 and Silver et al., 2003). Помимо формы, молекулярный состав опорно-двигательного аппарата также определяется функционально, посредством явления механотрансдукции (Ingber, 2006, Stevens-Tuttle et al., 2008 and Banes et al., 1999), которое будет рассмотрено далее.
В соответствии с вышеизложенным, повторяющиеся движения постепенно определяют форму и состав движущихся структур, а также связанные с ними пути нервной регуляции. Раннее появление и стабильность потягивания свидетельствуют о его вкладе в это развитие.
Нейрофармакология и нейрофизиология СРЗ: функция возбуждения?
СПЗ ранее связали с поддержанием уровня возбуждения ЦНС и внимания, т. е. он приводит центральную нервную и опорно-двигательную системы в такое состояние, а также поддерживает это состояние, в котором животное способно воспринимать стимулы из окружающей среды и реагировать на них соответствующими двигательными актами (Baenninger, 1997, Walusinski, 2006 and Askenasy, 1989).
Ни одна конкретная структура головного мозга не была ассоциирована с зеванием (Argiolas and Melis, 1998), поскольку зевание наблюдается также у новорожденных с анэнцефалией только при наличии продолговатого мозга (Heusner, 1946), однако, по-видимому, в процессе зевания также участвует ствол мозга. Кроме того, фармакологические и клинические данные свидетельствуют об участии гипоталамуса, луковицы и варолиева моста. Еще в 60-х гг. было показано, что инъекции различных веществ (например, адренокортикотропина (АКТГ), альфа-меланоцитостимулирующего гормона (a-МСГ) и связанных с ним пептидов), стимулируя деятельность ЦНС, вызывают активность СПЗ у лабораторных животных (Argiolas и Melis, 1998). Недавно было показано, что стереотипное зевание у крыс, включая потягивание туловища (СПЗ), может запускаться при стимуляции паравентрикулярного ядра (ПВЯ) гипоталамуса с помощью электрического или химического воздействия (веществами, которые выделяют окись азота) (Sato-Suzuki et al., 1998).
Сегодня считается, что после стимулирования ПВЯ окситоцинэргические нейроны выделяют окситоцин в экстрагипоталамические структуры и что эти структуры, в свою очередь, являются посредниками при реакциях СПЗ и возбуждения (Walusinski, 2006). Поскольку СПЗ включает различные поведенческие и автономные проявления, предполагается, что ПВЯ объединяет исходящую информацию из различных источников и создает эфферентные импульсы, по крайней мере, на пяти различных уровнях: респираторном, сердечно-сосудистом, лицевом двигательном, спинном двигательном и системы возбуждения (Sato-Suzuki et al., 1998), которые отвечают, соответственно, за реакцию глубокого дыхания, падение кровяного давления, открытие рта, потягивание туловища и реакцию возбуждения. Было показано, что другие нейротрансмиттеры, включая орексины, ацетилхолин, допамин, серотонин и опиоидные пептиды, также являются посредниками при реакции СПЗ (Argiolas and Melis, 1998 and Sato-Suzuki et al., 2002).
Модель Сато-Сузуки на реакции зевания/возбуждения у крыс демонстрирует основные пути ЦНС, которые, возможно, участвуют в СПЗ (Sato-Suzuki et al., 2002) (рис. 1).
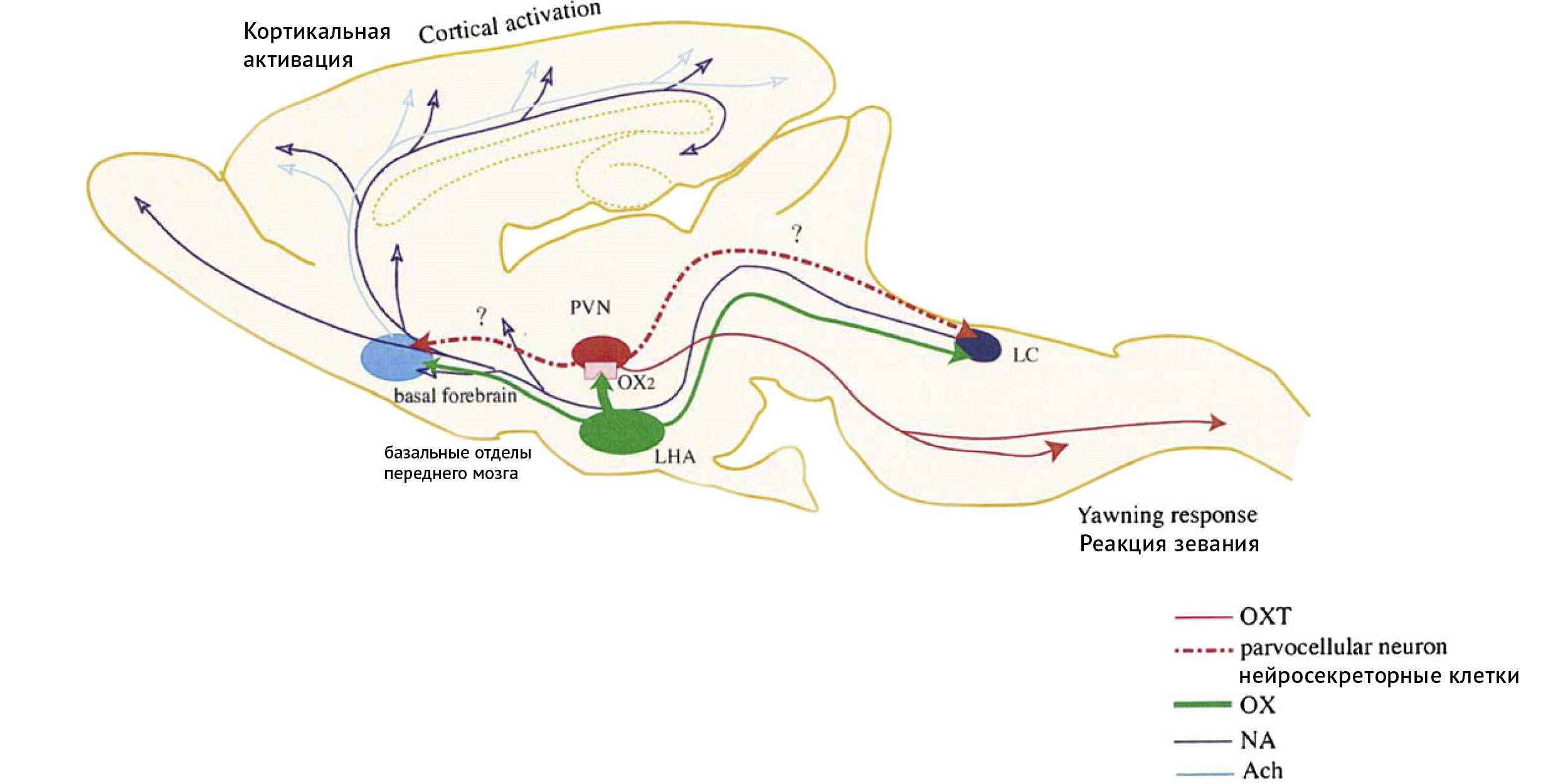
Рис. 1. Схематическое изображение модели Сато-Сузуки реакций зевания/возбуждения, которая осуществляется через ПВЯ у крыс. Нейроны орексина из латеральной гипоталамический области (ЛГО, LHA) могут прямо активировать нейроны в паравентрикулярном ядре (ПВЯ, PVN), которые, в свою очередь, проецируются в голубоватое пятно (LC) или базальные отделы переднего мозга. Нейроны окситоцина из ПВЯ проецируются в нижний ствол мозга, включая лицевое ядро и нейроны, связанные с дыхательной системой, которые задействованы в реакции зевания. OXT — окситоцин; OX — орексин; NA — норадреналин; Ach — ацетилхолин. Печатается с разрешения: Ikuko Sato-Suzuki, Ichiro Kita, Yoshinari Seki, Mitsugu Oguri, Hideho Arita. Behavioural Brain Research 128 (2002) 169-177 Cortical arousal induced by microinjection of orexins into the paraventricular nucleus of the rat.
Потягивание может рассматриваться как фиксированная форма действия (ФФД) (Rosenbloom, 1994) – упорядоченная последовательность рефлекторных действий. В эту категорию также входят такие эволюционно важные инстинктивные действия как питание, спаривание и другие виды социального поведения, имеющие важное значение для выживания. Потягивание, как представляется, вызывается сложным набором или последовательностью стимулов, которые могут включать как экстероцептивные сигналы (например свет — темнота), так и интероцептивные (например суточные эндокринные циклы и соматическая интероцепция) (Walusinski, 2006).
Фенотип СПЗ
СПЗ имеет схожий и стереотипный фенотип по всей эволюционной лестнице — он практически не изменился. Зевание (рис. 2) начинается с долгого и глубокого вдоха, достигает пика и завершается коротким выдохом. Дыхательные мышцы, мышцы рта, шеи и верхней части позвоночника совершают совместное сокращение, одновременно фиксируя суставы и растягивая миофасциальные ткани (Walusinski, 2006).

Рис. 2. Зевание обычно сопровождается потягиванием верхних конечностей.
Некоторые ссылки в существующей литературе описывают потягивание как серию скоординированных действий, которые разворачиваются последовательно, создавая сократительное натяжение мягких тканей до пиковой нагрузки, при котором стыки конечностей и туловища будут максимально расширены — или, в качестве альтернативы, туловище будет прогнуто в сгибании (Fraser, 1989b). После пика напряжение в мягких тканях резко падает, что и это вызывает ощущение удовольствия и благополучия (см. далее раздел, посвященный удовольствию). Эти действия могут охватывать отдельные области или все тело, и зачастую являются симметричными (Fraser, 1989b).
СПЗ: компенсаторная реакция на временную зажатость или неподвижность?
Паттерны потягивания всего тела в общем случае похожи на те, которые используются при ходьбе и выравнивании положения тела (Fraser, 1986b), т. е. они имитируют обычные функциональные движения, тогда как потягивание в отдельных областях тела, по-видимому, является корректирующей реакцией на зажатость, вызванную временным позиционным напряжением или неподвижностью.
В процессе метаболического круговорота компоненты внеклеточного матрикса (ВКМ) постоянно реабсорбируются и синтезируются. Молекулярные взаимодействия между глюкозоаминогликанами (ГАГ), волокнами и другими компонентами непрерывно изменяют структурно-механические свойства. Вплетение протоколлагеновых мономеров в волокна является спонтанным, оно происходит за счет гидрофильных взаимодействий водородных связей между водой и боковыми цепочками аминокислот. Все, что нужно для этого процесса, — это физиологические условия с определенной температурой, pH, ионной силой и степенью гидратации (Leikin et al., 1995 and Giraud-Guille et al., 2003).
Механические нагрузки являются основными факторами, определяющими форму и взаимосвязь в этом непрерывном и спонтанном переплетении, т. е. движение имеет решающее значение для поддержания надлежащей формы и функционирования системы ВКМ (Kjaer et al., 2005, Kjær et al., 2006, Kjær et al., 2009 и Tomiosso et al., 2005). Например, плотные ткани формируются в ответ на механическую потребность в сопротивлении натяжению, в то время как рыхлая соединительная ткань обновляется там, где требуется скольжение. Без механической нагрузки система ВКМ будет формироваться аморфно, а ее конфигурация не будет отвечать физиологическим потребностям.
Для того чтобы конфигурация системы ВКМ соответствовала физиологии, в первую очередь должна осуществляться соответствующая механическая нагрузка. Например, движение, обеспечивающее длинный шаг, будет оставаться возможным лишь в той мере, в которой оно достаточно выражено, поскольку только такое выражение будет стимулировать поддерживающую конфигурацию ВКМ (Kjær, 2004, Kjær et al., 2009 and Heinemeier et al., 2007).
Но в большинстве случаев животные не проявляют своих оптимальных качеств движения, таких как бег с максимальной скоростью и достижение максимальной амплитуды движения (АД) в суставах. Более того, сон создает регулярный период неподвижности, к которому адаптируются паттерны коллагеновых переплетений и взаимосвязей ВКМ. Это свидетельствует о непрерывной тенденции «связывать» всю структуру животного: перекрестные связи волокон коллагена, ориентированные в соответствии с неоптимальными движениями, могут ограничивать функцию путем уменьшения а) длины и прочности ткани и б) внутреннего скольжения между частями, необходимыми для достижения оптимального моторного потенциала. Если этому не противодействовать, то такой процесс может привести к постепенной утрате амплитуды движения в суставах и возникновению жесткости тканей. Эта этих фактора угрожают здоровью тканей, препятствуя распределению жидкости и искажая формы клеток, и, таким образом препятствуя корректной экспрессии генов. Ниже, в разделе, посвященном механотрансдукции, излагаются вопросы, касающиеся механической модуляции работы генов.
Потягивание, с характерной для него точной и энергичной работой мышц, может быть средством компенсации механических сигналов, поступающих в периоды отдыха и при неоптимальных движениях. В своих исследованиях потягивания у различных видов животных Фрэзер упоминает о том, что его можно рассматривать как обратную связь к неподвижности. Возможно также, что оно вызывается длительными периодами неподвижности в асимметричных положениях. Он приходит к выводу, что если тело имеет тенденцию к закрепощению, то потягивание «может служить для возвращения конечности (и связанных с ней мышц) в исходное (гомеостатическое) состояние» (Fraser, 1989a and Fraser, 1989b). Фактически СПЗ имеет много общего с другими гомеостатическими функциями, о чем говорится далее.
Эмоциональная двигательная система и СПЗ
Сознательная двигательная активность человека включает в себя согласованную деятельность всех составляющих двигательной системы. Запланированное движение переводится в реализуемые двигательные реакции через цепочку нейронной активности, которая начинается в префронтальной коре головного мозга, а затем последовательно достигает премоторной зоны и основной моторной коры головного мозга (Kandel et al., 2000 and Hallett, 2007). Аксоны нейронов первичной двигательной коры головного мозга, пролегая в корково-спинномозговом пути, передают импульсы непосредственно на двигательные нейроны. Ниже [основания продолговатого мозга – Прим. ред] эти аксоны образуют пирамиды, поэтому весь путь этих аксонов также называется пирамидальным. Двигательная информация, которую передает пирамидальный путь, модулируется не только за счет сенсорной информации, но и за счет информации, поступающей из моторных областей мозга, расположенных гораздо ниже (базальные ганглии, таламус, средний мозг, мозжечок, спинной мозг), и эта информация обеспечивает плавность, точность и точную координацию произвольных движений (Kandel et al., 2000). Модуляция информации на пирамидальном пути в основном является бессознательной и автоматической, т. е. подкорковые центры предоставляют бессознательные, непроизвольные и стереотипные компоненты, необходимые для управления движением (Jacobs and Horak, 2007, Guyton and Hall, 2006 and Takakusaki et al., 2003), в отличие от произвольных компонентов двигательных актов, которые управляются корковыми центрами.
В целом, чем ниже расположена структура, тем проще та моторная информация, которую она передает. Например, спинной мозг находится на самом низком уровне, и поступающая от него информация передает такие простые рефлексы как локомоторный рефлекс и рефлекс почесывания. Высшие центры обеспечивают выполнение более сложной двигательной активности, согласно иерархической организации сложности (Kandel et al., 2000). Например, повреждение первичной двигательной коры человека может привести к контрлатеральной гемиплегии, постуральные и стереотипные непроизвольные движения могут сохраняться (Kandel et al., 2000).
Аналогичным образом элементы инстинктивного поведения могут выполняться без вмешательства или модуляции со стороны корковых центров. Инстинктивное поведение развивались таким образом, чтобы гарантировать непрерывность наследственных линий организмов, поддерживать внутренний гомеостаз и обеспечивать успешное размножение (Dentona et al., 2009). Главным образом оно обеспечивается лимбической системой и включают в себя действия по поддержанию жизни (например, питание, самозащита, секс). Инстинктивное поведение собирательно именуются эмоциональным поведением (Guyton and Hall, 2006 и Kandel et al., 2000).
В экспериментальном моделировании на животных обнаружено, что стимуляция лимбических структур, в особенности гипоталамуса и миндалевидного тела, вызывает различные эмоциональные проявления (Kandel et al., 2000 and Guyton and Hall, 2006) даже при отсутствии коры головного мозга. Например, после декортикации –исследовательской процедуры, в ходе которой перерезается связь между корковыми и подкорковыми структурами – последние продолжают функционировать без тормозящего воздействия первых. Животные, подвергнувшиеся декортикации, могут питаться, выражать гнев, драться и совокупляться (Magoun and Ranson, 1938, Smith, 1939 and Guyton and Hall, 2006), а это показывает, что лимбическая система может обеспечивать такое поведение без участия коры головного мозга.
Фактически лимбическая система пронизывает бесчисленными диффузными аксонами продолговатый мозг. Эта система существует сама по себе, она способна производить двигательную активность независимо от произвольно управляемой пирамидальной системы. Хольтстейдж описывает ее как эмоциональную двигательную систему, потому что действия, которые регулируется из лимбической системы, зависят от эмоционального состояния животного. К функциям, регулируемым лимбической системой, относятся различные виды непроизвольных движений, связанных с обонянием и употреблением пищи, такие как лизание, жевание и глотание; клонические и ритмичные движения (например, локомоция, дрожь); сексуальные функции; использование голоса, смех и плач; оборонительные рефлексы и др. (Guyton and Hall, 2006 and Holstege, 1992). Действительно, у лабораторных животных воспроизводились различные стереотипные действия при электрической или химической стимуляции мезэнцефалических ядер, в частности ПВЯ (параветрикулярного ядра) гипоталамуса. Наблюдались жевание, лизание, лордоз у самок, эрекция полового члена и груминг (Argiolas et al., 2000, De Wied, 1999 and Vergoni et al., 1998). СПЗ входит в число тех видов действий, которые обеспечивает лимбическая система, и ее длительная филогенетическая устойчивость предполагает адаптивные функции.
Одна из функций ПВЯ заключается в интеграции нервных систем – центральной и периферической автономной. ПВЯ имеет контакт со всеми автономными нейронами в стволе мозга и спинном мозге, поскольку участвует в многочисленных автономных нервных и эндокринных функциях поддержания гомеостаза (Kandel et al., 2000). Эти функции включают: производство кортизола с помощью секреции кортикотропин-релизинг фактора (КРФ); выработку окситоцина и вазопрессина (гормонов, связанных с образованием молока и поддержанием водного баланса, соответственно); регулирование таких факторов, как потребление пищи, частота сердечных сокращений и кровяное давление (Kandel et al., 2000 and Holstege, 1992). То, что СПЗ запускается ПВЯ, предполагает, что СПЗ может относится к функциям общего гомеостаза. Эти функции осуществляются автономно эмоциональной двигательной системой.
Работу эмоциональной двигательной системы человека могут иллюстрировать непроизвольными движениями пациентов с повреждениями произвольных двигательных путей. Например, замечено, что при зевоте пациенты c гемиплегией непроизвольно поднимают парализованную руку (Graham, 1982, Töpper et al., 2003 and Stewart, 1921) и мышцы парализованной конечности активируются почти так же, как мышцы здоровой конечности при произвольном движении (Omam et al., 1989). Также у пациентов с гемиплегией возникают синкинезии (непроизвольные совместные движения), в которых произвольное перемещение одной части тела приводит к непроизвольному движению другой, например сгибание бедра и отведение руки во время чихания, или движения рук во время мочеиспускания (Walshe, 1923). Аналогичным образом пациенты с параличом лицевого нерва, при котором поражены нервные пути произвольных движений, способны демонстрировать лицевые движения при смехе (Hopf et al., 1992, Töpper et al., 1995 and Chernev et al., 2009) а пациенты, неспособные произвольным образом открывать рот, открывают его во время зевания (Askenasy, 1989). [На основании данного признака в англоязычной литературе параличи лицевого нерва подразделяют на Voluntary facial palsy и Emotional facial palscy – Прим. ред.]
В совокупности приведенные выше экспериментальные и клинические наблюдения иллюстрируют, что автоматическое (эмоциональное) двигательное поведение может выражаться в определенных паттернах независимо от произвольной соматомоторной системы. Поскольку у здоровых объектов первая система находится под тормозящим воздействием второй (Marder and Rehm, 2005 and Pincus and Tucker, 1974), непроизвольное лимбическое поведение обычно возникает при повреждении произвольных двигательных контуров – естественном (как при гемиплегии) или искусственном (как при декортикации). Тот факт, что обычное двигательное поведение человека включает модуляцию инстинктивных стимулов (Smith, 1992), может объяснить, почему СПЗ обычно снижается по мере старения человека.
Потягивание в сравнении с обычной растяжкой: автоматические и произвольные двигательные акты
Прислушавшись к своему интерцептивному чувству, можно уловить, что потягивание и СПЗ проявляют специфические качества двигательной системы. При намеренном «зевании», внутренние ощущения заметно отличаются от тех, которые появляются при спонтанном зевании. Аналогичным образом ощущения, возникающие при спонтанном потягивании, отличаются от тех, которые сопровождают либо «произвольное потягивание», либо произвольное растяжение мягких тканей.
Паттерны произвольной растяжки задаются когнитивно, а действие выполняется целенаправленно. Они часто включают расслабление мышц за счет уменьшения их рабочей нагрузки: мышца пассивно растягивается в результате либо силы тяжести, либо работы мышц-антагонистов.
В противоположность этому паттерны потягивания автоматические. В результате глубоких интенсивных и непроизвольных совместных мышечных сокращений мягкие ткани активно удлиняются относительно костных структур, поскольку суставы жестко закреплены. Каждое движение в рамках этого паттерна возникает последовательно, очевидно, благодаря вовлечению калейдоскопа рефлексов, последовательность которых не может быть ни предугадана, ни выполнена целенаправленно. Так же, как и спонтанное зевание по сравнению с намеренной имитацией зевания, спонтанное потягивание ощущается совсем по-другому по сравнению с произвольной растяжкой, лишь похожей на потягивание. Поскольку произвольные и эмоциональные двигательные системы имеют обособленные нейронные пути, отличительные внутренние ощущения, которые возникают при потягивании, могут быть отнесены к последовательностям вовлечения в процесс двигательных единиц, предназначенных для выполнения автоматических паттернов движения. Такое различие между интероцептивными ощущениями в ходе автоматических и произвольных двигательных актов и было зарегистрировано (Hommel, 2009). Более того, функционирование системы, имеющей высокий иерархический статус, может подавлять работу автоматической системы с более низким статусом . Это торможение может нарушать характерную спонтанность СПЗ в пользу когнитивно управляемого растяжения.
Автоматическое отведение руки во время зевания у пациентов с гемиплегией указывает на непроизвольную природу СПЗ: пациенты не воспроизводили движения руки, когда имитировали зевание (Töpper et al., 2003). Иными словами, сознательная имитация автоматического двигательного паттерна (через корково-спинномозговую систему) не приводит к немедленному воспроизведению инстинктивных паттернов, поступающих из лимбической системы (через эмоциональную двигательную систему). Кроме того, если модели двигательных паттернов различны, то их физиологические эффекты также должны отличаться.
Хорошо известно, насколько важна растяжка для поддержания здоровья опорно-двигательного аппарата. Каждый из множества адаптивных механизмов, обеспечивающих поддержание физической формы человека, включая растяжку, имеет свою причину существования. И хотя должны растягиваться все группы мышц, различные типы растяжений направлены на решение конкретных проблем и предназначены для компенсации различных паттернов укорочения мышц или последующего ограничения подвижности суставов. Но как животные в дикой природе поддерживают здоровье опорно-двигательного аппарата? Они не выполняют растяжку намеренно и все же сохраняют двигательные способности. Может ли СПЗ быть причиной этого?
Если это так и если было бы возможно стимулировать возникновение СПЗ, то можно ли использовать СПЗ с терапевтическими целями? Многочисленные практики психосоматической терапии включают СПЗ из-за его ярко выраженного гомеостатического воздействия, например в Соматике Ханны, практике Эутонии и Джойфлексинге (Hanna, 2004, Johnson, 2002 and Vishnivetz, 1995). В практике Эутонии СПЗ вызывают помощью определенных состояний внимания и форм механической стимуляции. Аналогичным образом специальная механическая стимуляция в репозиционировании мышц может также стимулировать СПЗ (см. ниже раздел, посвященный реакциям, вызванным РМ).
Удовольствие и здоровье
Древнее биологическое поведение, связанное с поддержанием гомеостаза, направляется через интерцепцию — сенсорные ощущения, отражающие физиологическое состояние (Craig, 2003). Сенсорные ощущения неудовольствия и удовольствия определяют эмоциональные качества стимулов, влияющих на поведение животного (Guyton and Hall, 2006 и Bozarth, 1994). Приятные ощущения, связанные с положительным влиянием, подкрепляют многие виды жизненно важных действий: удовлетворение голода и жажды, спаривание, а также опорожнение мочевого пузыря и кишечника являются — все это примеры инстинктивного поведения. Выполнив соответствующее действие, животное вознаграждается ощущением удовольствия, которое биологически подкрепляет удовлетворение потребности. Сохранение здоровья (салютогенез) неразрывно связано с позитивными чувствами (Esch and Stefano, 2004). Эти позитивные эмоциональные состояния тесно связаны с древними субнеокортикальными лимбическими областями мозга, которые являются общими для людей и других млекопитающих, у которых, как оказалось, гомеостатическое удовлетворение обеспечивается действием различных нейропептидов (Burgdorf and Panksepp, 2006, Vincent, 1994 and Cabanac, 1992).
Инстинктивное поведение, внося свой вклад в поддержание внутренней среды, может рассматриваться как поддержка гомеостаза (Sherwood, 2010) — категория, в которую также можно включить потягивание. Потягивание не только ассоциируется с удовольствием и благополучием (Fraser, 1989a, Sauer and Sauer, 1967, Russell and Fernández-Dols, 1997, Walusinski, 2006 and Stewart, 1921), но так же, как и другие факторы поддержки гомеостаза, задействует ПВЯ гипоталамуса. Гомеостаз в основном поддерживается парасимпатическим отделом автономной нервной системы (Recordati and Bellini, 2004), и во время проявления СПЗ обнаруживается повышенная парасимпатическая активность (Askenasy and Askenasy, 1996).
Более того, частота проявлений СПЗ связана с уровнями состояния здоровья и выздоровления: Фрейзер (Fraser, 1989b) отмечает, что потягивание отсутствует у животных с системными заболеваниями, но восстанавливается, когда животное выздоравливает. Аналогичным образом, у пациентов, восстанавливающихся после гемиплегии, вновь характерным образом появляются СПЗ и синкинезия [совместное рефлекторное движение – Прим. ред.] (Töpper et al., 2003 and Hwang et al., 2005) – вплоть до возобновления произвольного движения конечностей. Интересно, что паттерны непроизвольной синкинезии предложены в качестве параметра оценки восстановления после постинсультного гемипареза, поскольку сроки возникновения таких паттернов, по-видимому, связаны с функциональными результатами пациента (Hwang et al., 2005). Когда больной находится в состоянии комы, зевание предсказывает приближение момента выхода из нее (Braunwald et al., 1987). Терапевтическое применение транскраниальных электромагнитных полей для лечения болезни Паркинсона и рассеянного склероза вызывало СПЗ. В исследовании рассеянного склероза СПЗ снова появлялся у тех пациентов, болезнь которых, в ответ на лечение, перешла в стадию ремиссии (Sandyk, 1999 and Sandyk, 1998).
Однако чрезмерное потягивание связано с некоторыми болезнями и приемом ряда лекарств (Askenasy, 1989). Это заставляет предположить наличие возможного разграничения между полным (успешным) и неполным (неудачным) потягиванием. Возможно, что первый вид [потягивания] после выполнения своей задачи, возобновляется физиологически; тогда как второй повторяется патологически в стремлении к завершению. Рассмотрим, например, ощутимое разочарование, сопровождающее прерывание процесса зевания или чихания!
Механотрансдукция, тенсегрити и здоровье
Можно утверждать, что функционирование клетки существенно зависит от ее формы. Клетку более не рассматривают, как гелевую цитоплазму, окруженную гибкой мембраной. Теперь преобладает мнение, что структура клетки создается ее внутренним цитоскелетом, который состоит из сложной системы микротрубочек и микрофиламентов (Ingber, 2008a). Синтез цитоскелета, который определяет форму клетки, регулируется механическими стимулами (Patwari and Lee, 2008 and Bischofs et al., 2008). Более того, механические связи между цитоскелетом с одной стороны, и компонентами клетки, такими как аппарат синтеза белка (например рибосомы и мРНК) и мембранные рецепторы с другой стороны, механически модулируют функции компонентов с помощью механизма, известного как механотрансдукция (Ingber, 2008b, Bischofs et al., 2008 and Chiquet et al., 2009). При помощи механотрансдукции механические стимулы переводятся в химические реакции (Humphrey and Delange, 2004), такие как синтез белка. С помощью модуляции экспрессии генов, активности поверхностных мембранных рецепторов, ионных каналов и связанных с ними клеточных функций, механические силы управляют развитием, обслуживанием, функциями и всей судьбой клетки (например, Dahl et al., 2010, Chiquet et al., 2009, Patwari and Lee, 2008 and Haudenschild et al., 2009).
Форма клетки, ее молекулярный состав и функции отражают воздействующие на нее механические напряжения. Например, в соответствии с конкретными динамическими паттернами механической нагрузки, мезенхимальные стволовые клетки позволяют создавать отличительные паттерны экспрессии генов, которые, в свою очередь, создают изменения в форме и объеме клеток (Haudenschild et al., 2009). Как представляется, каждая ткань развивается и поддерживается для того, чтобы выдерживать те воздействия, которым она подвергается. Например, напряжение стимулирует гены как фибробластов, так и остеобластов, в то время как сжатие стимулирует гены, связанные с хондрогенезом (Haudenschild et al., 2009). Данные свидетельствуют о том, что определенные механические стимулы могут даже помочь клеткам с дефектами генома достигнуть нормального фенотипа (Gieni et Hendzel, 2008).
Однако поскольку клетки не являются изолированными, на форму клетки также влияет связь клетки с окружающим ее внеклеточным матриксом (ВКМ). Благодаря взаимодействию на адгезивных участках клеточной мембраны, компоненты системы ВКМ влияют на общий механический результат воздействия внешних сил (Streuli, 2009 and Bischofs et al., 2008). Например, фибробласты и другие адгезивные клетки ощущают физические параметры (например жесткость) ВКМ и меняют соответственно экспрессию своих белков, таким образом изменяя собственную морфологию (Döbereiner et al., 2005 and Chiquet et al., 2009).
Важно отметить, что благодаря воздействию ВКМ клетка ведет себя подобно жидкому кристаллу (Giraud-Guille et al., 2003, Giraud-Guille, 1992, Bouligand, 1972 and Kreis and Boesch, 1994). Таким образом, рост, форма и функция живых тканей, в отличие от макромолекулярных кристаллов, которые выращиваются реакторах, регулируются механическими, независимыми от биологической активности стимулами (Lappa, 2003). Таким образом, по-видимому, механические напряжения определяют состав и форму ВКМ (и, следовательно, форму и паттерны мобильности организма) как биологическим (с помощью механотрансдукции), так и небиологическим способами (благодаря его жидкостнокристаллическим свойствам).
На макроскопическом уровне ВКМ можно концептуально рассматривать как волокнистый скелет (Bienfait, 1987), связанный с костным скелетом. Эти два скелета, как считается, образуют структуру тенсегрити (Ingber, 1998 and Ingber, 2008): кости, сопротивляясь сжатию, удерживают дистанцию между суставами и противодействуют базальному натяжению (предварительному стрессу), которое вызывают мягкие ткани. [Тенесгрити — целостности структуры в ее натяжении; такое свойство каркасной структуры, обладающей элементами натяжения и сжатия, при котором каждый элемент системы работает с максимальной эффективностью и экономичностью. Термин изобретен Бакминистером Фуллером. Также «Тенесгрити» называется система движений, созданная Карлосом Кастаньедой (в этом смысле в статье не используется) — Прим. ред.] Мягкие ткани — это элементы натяжения, которые поддерживают относительные положения костей. Тенсегрити также существует на более мелких уровнях. Например, структурные иерархии связывают макроскопические соединения «костей» и «мягких тканей» в механочувствительный цитоскелет, который связан с клеточными компонентами, регулирующими экспрессию генов (Ingber, 2006). Движения и осанка — действуя одновременно на макроуровне и на микроуровне — механически регулируют функции клеток, их форму и соотношения в ВКМ (Kjær, 2004, Kjær et al., 2009 and Heinemeier et al., 2007).
Наблюдая за перемещением пешеходов на многолюдной улице, можно увидеть, насколько индивидуальны человеческие движения. Среди огромного разнообразия двигательных паттернов можно отметить качества движения: координированные, плавные и изящные движения у одних людей, и затрудненные, резкие и неловкие движения — у других. Не все качества движения передают стимулы, одинаково полезные для формы и функций клеток. На здоровье и болезнь человека оказывают влияние как положительные, так и отрицательные стрессы [дистрессы – Прим. ред.].
Возможно, оптимальное движение — это такое движение, которое правильно распределяет механическую нагрузку на различные ткани и клетки, стимулируя их в соответствии с их потребностями. Каждой клетке для нормального функционирования требуется особый механический стимул (Ingber, 2006), а структурные и механические характеристики ВКМ — это результирующие постоянно изменяющихся воздействующих на него механических нагрузок.
Короче говоря, механическое равновесие диктует распределение напряжений между твердыми и мягкими, а оно, в свою очередь, играет ключевую роль в том, какую форму примут клетки и каков будет метаболизм. При потягивании сильное совместное сокращение групп мышц-антагонистов создает интенсивные механические импульсы. Они могут выступать в роли соответствующих организующих сигналов для клеток и тканей за счет изменения механических условий, оптимальных для их окружающей среды.
Оптимальные механические импульсы могут обеспечивать потенциал для оптимальной двигательной функции
Энергичный механический импульс, возникающий при СПЗ, максимально растягивает мягкие ткани и разгибает суставы, что, в свою очередь, максимально увеличивает размеры тела. Например, зевание увеличивает диаметр глотки в четыре раза и разводит голосовые связки до степени, которая «не наблюдается ни в какой другой момент жизни» (Walusinski, 2006). СПЗ может быть биологической компенсацией за периоды неподвижности и/или неправильных положений тела, восстанавливая подвижность животного за счет разрывов анормальных перекрестных связей в ВКМ, образовавшихся во время бездействия или неоптимальной деятельности.
Аномальные перекрестные связи в ВКМ могут вызывать сокращение мягких тканей и уменьшать количество внутренней смазывающей жидкости, что приводит к уменьшению подвижности. СПЗ может восстанавливать длину и нормальное внутреннее скольжение мягких тканей. В то же время механотрансдукция поддерживает постоянство состава ткани, подстраивая компоненты ВКМ согласно механическим потребностям клеток и защищая их таким образом от механических повреждений (Banes et al., 1999 and Ingber, 2006).
СПЗ и состояние пробуждения: восстановление постурального тонуса и тенсегрити
Зевание чаще встречается у хищников, которые обладают ярко выраженными циклами покоя и активности, чем у травоядных, поведение которых демонстрирует более длительные циклы возбуждения (Baenninger, 1997). Это еще одно доказательство функции возбуждения, которую обеспечивает СПЗ. При пробуждении ретикулярная формация передает активирующие импульсы не только в таламическо-корковые системы, но также и в постуральную и локомоторную системы. Эта активация обеспечивает легкий доступ к реакции борьбы или бегства [адаптационная реакция переактивации – Прим. ред.] (Walusinski, 2006).
Тот факт, что животные наиболее часто пробуждаются сразу после БДГ-фазы [быстрых движений глаз — Прим. пер.] сна (которая характеризуется мышечной атонией), позволил Валюсински (Walusinski, 2006) постулировать противоположную взаимосвязь между БДГ-фазой сна и СПЗ. Согласно Валюсински, сильные мышечные сокращения вызывают резкий переход от нейронных паттернов, контролирующих БДГ-фазу сна, к паттернам, способствующим активации функциональной сети, которая соответствует управляемому двигательному паттерну бодрствования. Видимо, СПЗ восстанавливает в миофасциальной системе повышенный уровень тонуса, необходимый для деятельности в условиях воздействия силы тяжести: ведь во сне миофасции провисают и расслабляются, и после пробуждения их нужно снова «собрать», прежде чем организм сможет правильно двигаться в поле силы тяжести.
Для структуры, обладающей свойством тенесгрити характерно то, что упорядоченное распределение предварительно прикладываемого напряжения обеспечивает быструю механическую коммуникацию, которая охватывает все тело. Растянутые элементы организма обеспечивают импульсу возможность достичь механорецепторов и повлиять на них в любой части системы, вне зависимости от начальной точки приложения импульса и его направления. Таким образом, структура со свойством тенсегрити демонстрирует гораздо большую механочувствительность по сравнению с расслабленной системой, в которой влияние внешних стимулов остается локальным. В уравновешенной [внутренне] структуре тенсегрити нет провисания, а это означает, что механическая коммуникация происходит практически мгновенно. Это обеспечивает быстрые и точные двигательные реакции (Ingber, 2006, Ingber, 2008b and Chen and Ingber, 1999), адаптивное значение которых очевидно. Структурная связь между элементами в системе, обладающей свойством тенсегрити, называется морфологической коммуникацией. Выдвигалось предположение, что она, наряду с нервной системой, обеспечивает контроль движений (Rieffel et al., 2010).
Чтобы основное натяжение в структуре тенсегрити распределялось эффективно, связи между натяжными элементами должны быть расположен очень точно. Кроме того, элементы сжатия также должны быть сближены очень точно относительно друг друга, что требует свободной суставной амплитуды движения и свободного скольжения среди действующих вместе – и точно расположенных — миофасциальных структур. Для «сборки» костного и фиброзного скелетов, которые в полной мере проявят свойство тенесгрити, необходима функциональная комбинация связей между элементами натяжения, способными скользить между элементами сжатия. Каждая форма тела (особая организация межсегментных относительных положений) требует строго определенного сочетания точно расположенных структурных связей и конфигураций скольжения, которые обеспечивают равномерное распределение предварительного напряжения. Невероятное разнообразие форм, которые принимает тело, чтобы совершить определенное движение, на каждое из которых воздействует гравитация, предполагает, что число возможных конфигураций связей и скольжения бесчисленны.
Возможно, энергичные совместные сокращения [мышц] в процессе потягивания систематически меняют структурную связь между сегментами и одновременно дают клеткам сигнал (посредством механотрансдукции) синтезировать компоненты ВКМ, необходимые для поддержания соответствующей [внутренней] среды (рис. 3). Если это так, то потягивание может способствовать восстановлению оптимальных опорно-двигательных механизмов и таким образом оптимизировать двигательные функции.

Рис. 3. Собаки часто потягиваются, разгибая передние конечности и туловища, часто сопровождая потягивание зеванием. Потягивание включает последовательное совместное сокращение и растягивание мягких тканей, которое может захватывать практически на все мышцы тела.
Связь с потягиванием: йога и боевые искусства
Поза «собака мордой вниз» (рис. 4), как и многие асаны йоги, напоминает позицию потягивания животных (Iyengar, 1979). Существует мнение, что йога берет начало в автоматических и спонтанных движениях опытных йогинов, пребывавших в состоянии глубокой медитации, и что йога должна практиковаться спонтанно (Muni, 1994). Восточные боевые искусства также могут быть связаны с потягиванием. В цигун, например, тело укрепляется за счет автоматического (непроизвольного) тонуса глубоких постуральных мышц. При этом поверхностные мышцы, связанные с произвольными движениями, расслаблены (рис. 5).

Рис. 4. Асана йоги «собака мордой вниз» (Адхо Мукха Шванасана) напоминает потягивающуюся собаку.
В ходе выполнения практики тело действует как целое, а все его части связаны друг с другом в движении (см. http://www.caiwenyu.com.br/09_Fotos_p_ing.htm). [Здесь и далее автор, вероятно, путает цигун и тайцзы-цюань. Цигун не всегда задействует движения, основываясь в большей степени на дыхательных практиках, тогда как два упомянутых принципа является базовыми для тайцзы-цюань, которое берет начало из боевых искусств: «все тело должно быть цепью, в которой звенья — суставы» (Чжан Саньфэн «Тайцзицюань-лунь») — Прим. ред.] Невозможно реализовать эти принципы с помощью произвольных двигательных актов, однако они возникают спонтанно при состоянии внимания, которое усиливает чуткость к механическим импульсам. Человек, находясь в таком состоянии и выполняя толчок, может воспользоваться упругой потенциальной энергией, накопленной в организме. Эта особенность цигун предполагает режим движений, основанный с свойстве тенесгрити и высоким уровнем предварительного натяжения. Увеличение работоспособности предварительно растянутых мышц, благодаря их способности сохранять упругую потенциальную энергию, уже было отмечено в литературе (Bosco et al., 2008, Ettema et al., 1990 and Ishikawa et al, 2006). [Для обозначения способности увеличивать силу мышцы за счет сохраненной потенциальной энергии Автор использует термин «потенцирование» (potentiation). Потенцирование – увеличение силы нервного импульса вдоль проходящего пути, который прежде использовался для сокращения или растяжения мышцы. – Прим. ред. ]

Рис. 5. Поза цигун. Растяжение всей миофасциальной системы напоминает потягивание.
Элементы обучения боевым искусствам часто описываются в терминах, которые подразумевают паттерны потягивания животных (Johnson, 2002). [Вероятно, имеется ввиду практика «Пять зверей» цигун, основанная на паттернах животных движений в спокойном состоянии. «Звериные» стили боевых искусств не имеют отсылок к потягиванию. – Прим. ред. ] Это заставляет задуматься над тем, что кошки и собаки, которые подверглись декортикации, проявляют инстинктивное поведение, например едят, совокупляются и дерутся (Argyle, 1988). Иными словами, базовое жизнеобеспечивающее поведение может проявляться и без участия коры головного мозга. Фактически драки являются в значительной степени рефлекторным поведением, проявление которого связано с подкорковыми структурами, такими как гипоталамус и центральное серое вещество (ЦСВ) среднего мозга (Ulrich and Azrin, 1962 and Shaikh and Siegel, 1994).
Реакции, вызванные РМ, могут быть похожи на потягивание
Подобно потягиванию, мануальная местная нагрузка миофасциальной системы от РМ интегрирует части тела, вызывая, по-видимому, совместное сокращение противоположных групп мышц (Bertolucci, 2008, Bertolucci and Kozasa, 2010a and Bertolucci, 2010b). При этом она вызывает заметное повышение тонической мышечной активности, которая указывает на общее увеличение нагрузки. Субъективный опыт клиента аналогичен тому, который вызывает потягивание, что предполагает наличие общих элементов с йогой, боевыми искусствами и РМ.
При потягивании мышечная активация начинается локально и распространяется на соседние области, пока не достигнет пика распространения и интенсивности. Иными словами, благодаря цепи рефлексов суставы поочередно теряют подвижность. Затем в процесс вовлекаются соседние сегменты, чтобы последовательно сформировать еще более крупный блок, который в конечном счете охватывает все тело. После пика [напряжения] ткани расслабляются.
РМ вызывает аналогичное последовательное вовлечение сегментов тела. Включение каждого сегмента увеличивает общее напряжение в блоке пока, вслед за пиком [напряжения], специалист не ощутит резкое расслабление мягких тканей. Последовательное вовлечение сегментов в процесс сопровождается усиливающейся непроизвольной активностью тонических мышц, которую можно обнаружить как путем пальпации, так и с помощью электромиографии (Bertolucci, 2008, Bertolucci and Kozasa, 2010a and Bertolucci, 2010b).
Давайте представим, каким образом РМ и потягивание могут вызывать схожую активность мышц. Автор выдвигает гипотезу о том, что приложенная в ходе РМ сила рук, имитирует внутренние силы и, таким образом, стимулирует афферентные механорецепторов с характеристиками, подобными тем, что возникают при потягивании.
В клинических условиях у людей, проходящих процедуры РМ, наблюдались спонтанные движения, похожие на потягивание (см. видео: http://musclerepositioning.blogspot.com/), и они описывали свои субъективные ощущения в ходе проведения РМ похожими на ощущения при потягивании. Некоторые клиенты также сообщали, что возобновили привычку потягиваться по утрам, считая, что благодаря ей они глубже ощущают телесное здоровье и что она помогает им облегчить проявление [болезненных] симпотомов опорно-двигательного аппарата. Эти наблюдения поддерживают гипотезу о сходстве между РМ и потягиванием.
Возможно, РМ представляет собой смесь миофасциального расслабления и «сопровождаемого потягивания», в ходе которой расслабление мягких тканей вызывает сочетание работы рук специалиста и тонических реакций, подобных потягиванию, генерируемых за счет внутренних сил. Эта комбинация сил может более эффективно воздействовать на мягкие ткани, чем манипуляции руками и потягивание по отдельности.
Подавляется ли СПЗ культурными традициями?
СПЗ задействует генетически запрограммированный центральный генератор упорядоченной активности продолговатого мозга, которые у высших приматов находятся под контролем неокортекса (Marder and Rehm, 2005). [Центральный генератор упорядоченной активности – распределенная сеть, только некоторые ее элементы локализуются в продолговатом мозге. – Прим. ред]. Отмечено, что люди, в отличие от других приматов, по мере старения зевают реже (Walusinski et al., 2005). Однако, у разных видов обезьян из Старого Света (Cercocebus albigena, Macaca fuscata и Macaca fascicularis) зевание становится все более частым по мере повышения уровня андрогенов в плазме крови (Deputte, 1994 and Troisi et al., 1990).
Поскольку зевание имеет негативное социальное значение в большинстве культур и религиозных традиций (Walusinski et al., 2010), возможно, культурная обусловленность подавляет СПЗ у людей. Учитывая вероятную гомеостатическую функцию СПЗ, любое такое подавление может способствовать повышению количества заболеваний опорно-двигательного аппарата у людей.
Биологические часы и здоровье
Поддержание гомеостаза тесно связано с биологическими ритмами, которые контролируются как внешними, так и внутренними стимулами (Aréchiga, 2003). Биологические ритмы формируются за счет внутриклеточных механизмов, которые называются циркадными часами. Они есть в обоих специальных центрах ЦНС и в большей части периферических тканей (Duguay and Cermakian, 2009 and Aréchiga, 2003). В частности, они присутствуют в фибробластах — клетках, которые отвечают за синтез компонентов ВКМ. Было показано, что внутренние биологические ритмы фибробластов участвуют в различных метаболических функциях (например в синтезе белков цитоскелета, контроле клеточного цикла, синтезе ферментов и др.) (Bursian, 2009). Хотя метаболический механизм фибробластов имеет собственный ритм, на него также влияют механические раздражители, в т. ч. предположительно те, которые генерируются за счет других биологических ритмов. Это ставит вопрос влияют ли друг на друга разные биологические ритмы.
Известно, что различные биологические ритмы связаны друг с другом. Например, между собой взаимодействуют частота сердечных сокращений, дыхание, периодические и соматодвигательные возбуждения (De Haro and Panda, 2006Niizeki et al., 1993). Существование таких сложных взаимоотношений между биологическими ритмами привело к концепции мультиколебательных систем, а также к гипотезе о том, что синхронизация различных сосуществующих ритмов является определяющим фактором экспрессии генов и биологической активности как у растений, так и у животных (Lloyd, 2009 and Bell-Pedersen et al., 2005). Вполне возможно, что циркадные часы фибробластов могут регулярно коррелировать с другими биоциклическими явлениями, такими, например, как связанный с СПЗ переход сон-бодрствование.
Было показано, что ритм сна и бодрствования и другие биологические ритмы связаны с иммунным ответом (Shepard and Shek, 1997), что указывает на их важное значение для общего состояния здоровья. Это вдохновляет на дальнейшее изучение возможной роли ритмических биологических процессов (таких как СПЗ) в поддержании гомеостаза как определенных тканей, так и организма в целом. Микродиализ тканей, метки метаболизма в сыворотке крови, нейровизуализация, ЭЭГ и ЭМГ — все это возможные способы дальнейшего изучения физиологической роли СПЗ.
Заключение
Концепция передачи миофасциальной силы (Huijing and Jaspers, 2005) предполагает наличие связей ВКМ среди костно-мышечных компонентов. Именно эти связи объединяют их в интегрированную систему, т. е. считается, что интегрирующую роль играют сами фасции. Интегрированное движение и требует, и стимулирует соответствующие связи в матрице. Однако животные совершают много неоптимальных движений, причинами которых являются неподвижность (например, во время сна), травмы и плохие постуральные привычки.
Если нормальная деятельность создает как «хорошие», так и «плохие» механические сигналы, поступающие в ВКМ, то, для того чтобы животное могло поддерживать возможность полноценного движения на протяжении всей жизни, плохие сигналы должны перекрываться хорошими. Потягивание может послужить одним из источников хороших сигналов за счет а) разрыва плохих связей при стимулировании хороших и б) восстановления тонуса мышц для осуществления интегрированного движения, которое является еще одним источником хороших механических сигналов. Короче говоря, потягивание может быть формой нейромиофасциальной гигиены.
Если это так, можно ли рекомендовать практиковать потягивание для улучшения общего состояния здоровья? Такая мера потребует переоценки культурного остракизма в отношении зевания и потягивания, а также дальнейшего изучения терапевтических подходов, подобных репозиционированию мышц, которые, по-видимому, стимулируют потягивание.
Благодарности
Я благодарен Йеде Бокалетто (Yeda Bocaletto), Анжеле Лобо (Angela Lobo), Сорайе Паккиони (Soraia Pacchioni) и Хейди Масса (Heidi Massa) за выход в свет данной рукописи.
Список использованной литературы
Aréchiga, H., 2003. The neural substrate of biological rhythms. Revista de Neurologia 36 (1), 49e60.
Argiolas, A., Melis, M.R., 1998. The neuropharmacology of yawning. European Journal of Pharmacology 343 (1), 1e16. doi:10.1016/S0014-2999(97)01538-0.
Argiolas, A., Melis, M.R., Murgia, S., Schio¨th, H.B., 2000. ACTH- and alpha-MSH-induced grooming, stretching, yawning and penile erection in male rats: site of action in the brain and role of melanocortin receptors. Brain Research Bulletin 51 (5), 425e431.
Argyle, M., 1988. Bodily Communication. Routledge, New York.
Askenasy, J.J., 1989. Is yawing an arousal defense reflex? The Journal of Psychology 123 (6), 609e621.
Askenasy, J.J., Askenasy, N., 1996. Inhibition of muscle sympathetic nerve activity during yawning. Clinical Autonomic Research 6, 237e239. doi:10.1007/BF02291140.
Baenninger, R., 1997. On yawning and its functions. Psychonomic Bulletin and Review 4 (2), 198e207.
Banes, A.J., Horesovsky, G., Larson, C., Tsuzaki, M., Judex, S., Archambault, J., Zernicke, R., Herzog, W., Kelley, S., Miller, L., 1999. Mechanical load stimulates expression of novel genes in vivo and in vitro in avian flexor tendon cells. Osteoarthritis Cartilage 7 (1), 141e153. doi:10.1053/joca.1998.0169.
Bell-Pedersen, D., Cassone, V.M., Earnest, D.J., Golden, S.S., Hardin, P.E., Thomas, T.L., Zora, M.J., 2005. Circadian rhythms from multiple oscillators: lessons from diverse organisms. Nature Reviews Genetics 6 (7), 544e556. doi:10.1038/nrg1633.
Bertolini, A., Gessa, G.L., 1981. Behavioral effects of ACTH and MSH peptides. Journal of Endocrinological Investigation 4, 241e251.
Bertolucci, L.F., 2008. Muscle Repositioning: “A new verifiable approach to neuro-myofascial release?”. Journal of Bodywork and Movement Therapies 12, 213e224. doi:10.1016/j.jbmt.2008.05.002.
Bertolucci, L.F., 2010b. Muscle Repositioning: combining subjective and objective feedbacks in the teaching and practice of a reflex-based myofascial release technique. International Journal of Therapeutic Massage and Bodywork 3 (1), 26e35.
Bertolucci, L.F., Kozasa, E.H., 2010a. Sustained manual loading of the fascial system can evoke tonic reactions: preliminary results. International Journal of Therapeutic Massage and Bodywork 3 (1), 12e14.
Bienfait, M., 1987. Bases physiologiques de la the´rapie manuelle. S.E.D. “Le Pousoe´”, Saint-Mont.
Bischofs, I.B., Klein, F., Lehnert, D., Bastmeyer, M., Schwarz, U.S., 2008. Filamentous network mechanics and active contractility determine cell and tissue shape. Biophysical Journal 95 (7), 3488e3496. doi:10.1529/biophysj.108.134296.
Bosco, C., Komi, P.V., Ito, A., 2008. Prestretch potentiation of human skeletal muscle during ballistic movement. Acta Physiologica Scandinavica 111 (2), 135e140. doi:10.1111/j.1748-1716.1981.
Bouligand, Y., 1972. Twisted fibrous arrangement in biological materials and cholesteric mesophases. Tissue and Cell 4 (2), 189e217. doi:10.1016/S0040-8166(72)80042-9.
Bozarth, M.A., 1994. Pleasure systems in the brain. In: Warburton, D.M. (Ed.), Pleasure: The politics and the reality. John Wiley & Sons, New York, pp. 5e14.
Braunwald, E., Isselbacher, K.J., Petersdorf, R.G., Wilson, J.D., Martin, J.B., Fauci, A.S., 1987. Harrison’s Principles of Internal Medicine. McGraw-Hill, New York.
Briscoe, J., Wilkinson, D.G., 2004. Establishing neuronal circuitry: hox genes make the connection. Genes and Development 18 (14), 1643e1648. doi:10.1101/gad.1227004.
Burgdorf, J., Panksepp, J., 2006. The neurobiology of positive emotions. Neuroscience Biobehavioral Reviews 30 (2), 173e187. doi:10.1016/j.neubiorev.2005.06.001.
Bursian, A.V., 2009. Organization of endogenous rhythms of motor functions. Journal of Evolutionary Biochemistry and Physiology 45 (6), 663e669. doi:10.1134/S0022093009060039. Cabanac, M., 1992. Pleasure: the common currency. Journal of Theoretical Biology 155 (2), 173e200.
Chen, C.S., Ingber, D.E., 1999. Tensegrity and mechanoregulation: from skeleton to cytoskeleton. Osteoarthritis and Cartilage 7 (1), 81e94. doi:10.1053/joca.1998.0164. Chernev, I., Petrea, R.E., Reynolds, M.S., Wang, F., 2009. The classical type of Foix-Chavany-Marie syndrome: assessment and treatment of dysphagia. The Internet Journal of Neurology 11 (1).
Chiquet, M., Gelman, L., Lutz, R., Maier, S., 2009. From mechanotransduction to extracellular matrix gene expression in fibroblasts. Biochimica and Biophysica Acta 1793 (5), 911e920. doi:10.1016/j.bbamcr.2009.01.012.
Craig, A.D., 2003. Interoception: the sense of the physiological condition of the body. Current Opinion in Neurobiology 13 (4), 500e505. doi:10.1016/S0959-4388(03)00090-4.
Dahl, K.N., Booth-Gauthier, E.A., Ladoux, B., 2010. In the middle of it all: mutual mechanical regulation between the nucleus and the cytoskeleton. Journal of Biomechanics 43 (1), 2e8. doi: 10.1016/j.jbiomech.2009.09.002.
De Haro, L., Panda, S., 2006. Systems biology of circadian rhythms: an outlook. Journal of Biological Rhythms 21 (6), 507e518. doi: 10.1177/0748730406294767.
de Vries, J.I., Visser, G.H., Prechtl, H.F., 1982. The emergence of fetal behavior: I. Qualitative Aspects Early Human Development 7 (4), 301e322. doi:10.1016/0378-3782(82)90033-0.
De Wied, D., 1999. Behavioral pharmacology of neuropeptides related to melanocortins and the neurohypophyseal hormones. European Journal of Pharmacology 375 (1e3), 1e11. doi: 10.1016/S0014-2999(99)00339-8.
Dentona, D.A., McKinleyc, M.J., Farrell, M., Egan, G.F., 2009. The role of primordial emotions in the evolutionary origin of consciousness. Consciousness and Cognition 18 (2), 500e514. doi:10.1016/j.concog.2008.06.009.
Deputte, B.L., 1994. Ethological study of yawning in primates. I: Quantitative analysis and study of causation in two species of old world monkeys (Cercocebus albigena and Macaca fascicularis). Ethology 98 (3e4), 221e245.
Do¨bereiner, H.G., Dubin-Thaler, B.J., Giannone, G., Sheetz, M.P., 2005. Force sensing and generation in cell phases: analyses of complex functions. Journal of Applied Physiology 98, 1542e1546. doi:10.1152/japplphysiol.01181.2004.
Duguay, D., Cermakian, N., 2009. The crosstalk between physiology and circadian clock proteins. Chronobiology International 26 (8), 1479e1513.
Esch, T., Stefano, G.B., 2004. The neurobiology of pleasure, reward processes, addiction and their health implications. Neuroendocrinology Letters 4 (25), 235e251.
Ettema, G.J.C., Van Soest, A.J., Huijing, P.A., 1990. The role of series elastic structures in prestretch-induced work enhancement during isotonic and isokinetic contractions. Journal of Experimental Biology 154, 121e136.
Fraser, A.F., 1989a. Pandiculation: the comparative phenomenon of systematic stretching. Applied Animal Behaviour Science 23, 263e268. doi:10.1016/0168-1591(89)90117-2.
Fraser, A.F., 1989b. The phenomenon of pandiculation in the kinetic behaviour of the sheep fetus. Applied Animal Behaviour Science 24 (2), 169e182. doi:10.1016/0168-1591(89)90044-0.
Gieni, R.S., Hendzel, M.J., 2008. Mechanotransduction from the ECM to the genome: are the pieces now in place? Journal of Cellular
Biochemistry 104 (6), 1964e1987. doi:10.1002/jcb.21364. Giraud-Guille, M.M., 1992. Liquid crystallinity in condensed type I collagen solutions. A clue to the packing of collagen in extracellular matrices. Journal of Molecular Biology 224 (3), 861e873.
Giraud-Guille, M.M., Besseau, L., Martin, R., 2003. Liquid crystalline assemblies of collagen in bone and in vitro systems. Journal of Biomechanics 36 (10), 1571e1579. doi:10.1016/S0021-9290(03)00134-9.
Graham, M., 1982. Assoctiated reactions in the hemiplegic arm. Scandinavian Journal of Rehabilitation Medicine 14 (3), 117e120.
Guyton, A.C., Hall, J.E., 2006. Textbook of Medical Physiology. Elsevier, Philadelphia.
Hallett, M., 2007. Volitional control of movement: the physiology of free will. Clinical Neurophysiology 118 (6), 1179e1192. doi: 10.1016/j.clinph.2007.03.019.
Hanna, T., 2004. Somatics: Reawakening the Mind’s Control of Movement, Flexibility, and Health. Da Capo Press, Cambridge.
Haudenschild, A.K., Hsieh, A.H., Kapila, S., Lotz, J.C., 2009. Pressure and distortion regulate human mesenchymal stem cell gene expression. Annals Biomedical Engeneering 37 (3), 492e502. doi:10.1007/s10439-008-9629-2.
Heinemeier, K.M., Olesen, J.L., Haddad, F., Langberg, H., Kjaer, M., Baldwin, K.M., Schjerling, P., 2007. Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types. The Journal of Physiology 582 (3), 1303e1316. doi:10.1113/jphysiol.2007.127639.
Heusner, A.P., 1946. Yawning and associated phenomena. Physiological Reviews 26, 156e168.
Holstege, G., 1992. The emotional motor system. European Journal of Morphology 30 (1), 67e79.
Hommel, B., 2009. Action control according to TEC (theory of event coding). Psychological Research 73 (4), 512e526. doi: 10.1007/s00426-009-0234-2.
Hopf, H.C., Mu¨ller-Forell, W., Hopf, N.J., 1992. Localization of emotional and volitional facial paresis. Neurology 42 (10), 1918e1923.
Huijing, P.A., Jaspers, R.T., 2005. Adaptation of muscle size and myofascial force transmission: a review and some new experimental results. Scandinavian Journal of Medicine and Science in Sports 15 (6), 349e380. doi:10.1111/j.1600-0838.2005.00457.x.
Humphrey, J.D., Delange, S.L., 2004. An Introduction to Biomechanics: Solids and Fluids, Analysis and Design. Springer,NewYork.
Hwang, I.S., Tung, L.C., Yang, J.F., Chen, Y.C., Yeh, C.Y., Wang, C.H., 2005. Electromyographic analyses of global synkinesis in the paretic upper limb after stroke. Physical Therapy 85 (8), 755e765.
Ingber, D.E., 1998. The architecture of life. Scientific American 278 (1), 48e57.
Ingber, D.E., 2006. Cellular mechanotransduction: putting all the pieces together again. FASEB e Federation of American Societies for Experimental Biology Journal 20 (7), 811e827.
Ingber, D.E., 2008a. Tensegrity and mechanotransduction. Journal of Bodywork and Movement Therapies 12 (3), 198e200. doi: 10.1016/j.jbmt.2008.04.038.
Ingber, D.E., 2008b. Tensegrity-based mechanosensing from macro to micro. Progress in Biophysics and Molecular Biology 97 (2e3), 163e179. doi:10.1016/j.pbiomolbio.2008.02.005.
Ishikawa, M., Komi, P.V., Finni, T., Kuitunen, S., 2006. Contribution of the tendinous tissue to force enhancement during stretcheshortening cycle exercise depends on the prestretch and concentric phase intensities. Journal of Electromyography and Kinesiology 16 (5), 423e431.
Iyengar, B.K.S., 1979. Light on Yoga. Schocken Books, New York.
Jacobs, J.V., Horak, F.B., 2007. Cortical control of postural responses. Journal of Neural Transmission 114 (10), 1339e1348. doi:10.1007/s00702-007-0657-0.
Johnson, R., 2002. Boa forma para preguic¸osos: Joyflexing. Editora Pensamento-Cultrix, Sa˜o Paulo.
Jones, D.A., Round, J.,Haan, A., 2004. SkeletalMuscle from Molecules to Movement. A Textbook of Muscle Physiology for Sport, Exercise, Physiotherapy and Medicine. Churchill Livingstone, London.
Kandel, E.R., Schwartz, J.H., Jessell, T.M., 2000. Principles of Neural Science. McGraw-Hill, New York.
Kapandji, I.A., 1987. Fisiologia articular: Esquemas comentados de mecaˆnica humana. Ed Manole, Sa˜o Paulo.
Kjær, M., 2004. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiological Reviews 84 (2), 649e698. doi:10.1152/physrev.00031.2003.
Kjær, M., Langberg, H., Heinemeier, K., Bayer, M.L., Hansen, M., Holm, L., Doessing, S., Kongsgaard, M., Krogsgaard, M.R., Magnusson, S.P., 2009. From mechanical loading to collagen synthesis, structural changes and function in human tendon. Scandinavian Journal of Medicine and Science in Sports 19 (4), 500e510.
Kjaer, M., Langberg, H., Miller, B.F., Boushel, R., Crameri, R., Koskinen, S., Heinemeier, K., Olesen, J.L., Døssing, S., Hansen, M., Pedersen, S.G., Rennie, M.J., Magnusson, P., 2005. Metabolic activity and collagen turnover in human tendon in response to physical activity. Journal of Musculoskeletal and Neuronal Interactions 5 (1), 41e52.
Kjær, M., Magnusson, P., Krogsgaard, M., Boysen Møller, J., Olesen, J., Heinemeier, K., Hansen, M., Haraldsson, B., Koskinen, S., Esmarck, B., Langberg, H., 2006. Extracellular matrix adaptation of tendon and skeletal muscle to exercise. Journal of Anatomy 208 (4), 445e450.
Kreis, R., Boesch, C., 1994. Liquid-crystal-like structure of human muscle demonstrated by in vivo observation of direct dipolar coupling in localized proton magnetic resonance spectroscopy. Journal of Magnetic Resonance B 10 (2), 189e192. 4.
Lagercrantz, H., Ringstedt, T., 2001. Organization of the neuronal circuits in the central nervous system during development. Acta Paediatrica 90 (7), 707e715.
Lappa, M., 2003. The growth and the fluid dynamics of protein crystals and soft organic tissues: models and simulations, similarities and differences. Journal of Theoretical Biology 224 (2), 225e240.
Lehmann, H.E., 1979. Yawning: a homeostatic reflex and its psychological significance. Bulletin of the Menninger Clinic 43 (2), 123e136.
Leikin, S., Rau, D.C., Parsegian, V.A., 1995. Temperature-favoured assembly of collagen is driven by hydrophilic not hydrophobic interactions. Nature Structural Biology 2 (3), 205e210. doi: 10.1038/nsb0395-205.
Lloyd, D., 2009. Oscillations, synchrony and deterministic chaos. Progress in Botany 70, 69e91. doi:10.1007/978-3-540-68421-3_4.
Magoun, H.W., Ranson, S.W., 1938. The behavior of cats following
Bilateral Removal of the Rostral Portion of the cerebral Hemispheres. Journal of Neurophysiology 1, 39e44.
Marder, E., Rehm, K.J., 2005. Development of central pattern generating circuits. Current Opinion in Neurobiology 15 (1), 86e93. doi:10.1016/j.conb.2005.01.011.
Moore, S.W., 2003. Scrambled eggs: mechanical forces as ecological factors in early development. Evolution and Development 5 (1), 61e66. doi:10.1046/j.1525-142X.2003.03010.x.
Muni, S.W., 1994. Awakening the life force: The philosophy and psychology of “spontaneous yoga”. Llewellyn Publications, Minnesota.
Niizeki, K., Kawahara, K., Miyamoto, Y., 1993. Interaction among cardiac, respiratory, and locomotor rhythms during cardiolocomotor synchronization. Journal of Applied Physiology 75 (4), 1815e1821.
Oman, R.E., Sullivan, S.J., Fleury, J., Dutil, E., 1989. Yawning: a possible Confounding Variable in EMG Biofeedback studies
Biofeedback and self-Regulation. Applied Psychophysiology and Biofeedback 14 (4), 339e346. doi:10.1007/BF00999125.
Patwari, P., Lee, R.T., 2008. Mechanical control of tissue morphogenesis. Circulation Research 103 (3), 234e243. doi: 10.1161/CIRCRESAHA.108.175331. Pincus, J.H., Tucker, G.J., 1974. Behavioral Neurology. Oxford University Press, New York.
Recordati, G., Bellini, T.G., 2004. A definition of internal constancy and homeostasis in the context of non-equilibrium thermodynamics. Experimental Physiology 89 (1), 27e38. doi:10.1113/ expphysiol.2003.002633.
Rieffel, J.A., Valero-Cuevas, F.J., Lipson, H., 2010. Morphological communication: exploiting coupled dynamics in a complex mechanical structure to achieve locomotion. Journal of the Royal Society Interface 7, 613e621. doi:10.1098/rsif.2009.0240.
Rosenbloom, L., 1994. Motor development in early and later childhood: longitudinal approaches. Archieves of Disease Childhood 71 (4), 391.
Russell, J., Ferna´ndez-Dols, J.M., 1997. The Psychology of Facial Expression. Cambridge University Press, Cambridge, UK.
Sadler, T.W., 1995. Langman’s Medical Embriology, seventh ed.. Williams & Wilkins, Baltimore.
Sandyk, R., 1998. Yawning and stretching-a behavioral syndrome associated with transcranial application of electromagnetic fields in multiple sclerosis. International Journal of Neuroscience 95 (1e2), 107e113. doi:10.3109/00207459809000654.
Sandyk, R., 1999. Yawning and stretching induced by transcranial application of AC pulsed electromagnetic fields in Parkinson’s disease. International Journal of Neuroscience 97 (1e2), 139e145. doi:10.3109/00207459908994308.
Sato-Suzuki, I., Kita, I., Oguri, M., Arita, H., 1998. Stereotyped yawning responses induced by electrical and chemical stimulation of paraventricular nucleus of the rat. Journal of Neurophysiology 80 (5), 2765e2775.
Sato-Suzuki, I., Kita, I., Seki, Y., Oguri, M., Arita, H., 2002. Cortical arousal induced by microinjection of orexins into the paraventricular nucleus of the rat. Behavioral Brain Research 128 (2), 169e177. doi:10.1016/S0166-4328(01)00307-2.
Sauer, E.G., Sauer, E.M., 1967. Yawning and other maintenance activities in the South African Ostrich. The Auk 84, 571e587.
Shaikh, M.B., Siegel, A., 1994. Neuroanatomical and neurochemical mechanisms underlying amygdaloid control of defensive rage behavior in the cat. Brazilian Journal of Medical and Biological Research 27 (12), 2759e2779. Dec.
Shephard, R.J., Shek, P.N., 1997. Interactions between sleep, other body rhythms, immune responses, and exercise. Canadian Journal of Applied Physiology 22 (2), 95e116.
Sherwood, L., 2010. Human Physiology: From Cells to Systems, seventh ed.. Brooks/Cole, Belmont CA.
Silver, F.H., Siperko, L.M., Seehra, G.P., 2003. Mechanobiology of force transduction in dermal tissue. Skin Research and Technology 9 (1), 3e23. doi:10.1034/j.1600-0846.2003.00358.x.
Smith, K.U., 1939. The behavior of decorticate guinea pigs. Journal of Comparative Psychology 27 (3), 433e447.
Smith, R., 1992. Inhibition: History and Meaning in the Sciences of Mind and Brain. University of Califo´rnia Press, Berkeley.
Stevens-Tuttle, D., Fox, J., Bouffard, N.A., Henry, S., Wu, J., Langevin, H.M., 2008. Perimuscular fascia remodeling in a Porcine movement restriction model relevant to human low back pain. Journal of Bodywork and Movement Therapies 13 (1), 91. doi:10.1016/j.jbmt.2008.04.011.
Stewart, J.P., 1921. On muscle-tonus, tonic rigidity, and tonic Fits. British Medical Journal 1 (3137), 217e219.
Streuli, C.H., 2009. Integrins and cell-fate determination. Journal of Cell Science 122 (2), 171e177. doi:10.1242/10.1242/jcs.018945.
Takakusaki, K., Ohinata-Sugimoto, J., Saitoh, K., Habaguchi, T., 2003. Role of basal ganglia-brainstem systems in the control of postural muscle tone and locomotion. Progresss in Brain Research 143, 231e237.
Tomiosso, T.C., Gomes, L., de Campos Vidal, B., Pimentel, E.R., 2005. Extracellular matrix of ostrich articular cartilage. Biocell 29 (1), 47e54.
Topper, R., Kosinski, C., Mull, M., 1995. Volitional type of facial palsy associated with pontine ischaemia. Journal of Neurology. Neurosurgery and Psychiatry 58 (6), 732e734. doi:10.1136/jnnp.58.6.732.
Topper, R., Mull, M., Nacimiento, W., 2003. Involuntary stretching during yawning in patients with pyramidal tract lesions: further evidence for the existence of an independent emotional motor system. European Journal of Neurology 10 (5), 495e499. doi: 10.1046/j.1468-1331.2003.00599.x.
Troisi, A., Aureli, F., Schino, G., Rinaldi, F., De Angeli, N., 1990. The influence of age, sex, rank on yawning behavior in two species of macaques (Macaca fascicularis, Macaca fuscata). Ethology 86, 303e310.
Ulrich, R.E., Azrin, N.H., 1962. Reflexive fighting in response to aversive stimulation1 Journal of Experimental Analysis of Behavior. October 5 (4), 511e520. doi:10.1901/jeab.1962.5-511. Urba-Holmgren, R., Gonzalez, R.M., Holmgren, B., 1977. Is yawning cholinergic response? Nature 267 (5608), 261e262. doi: 10.1038/267261a0.
Vergoni, A.V., Bertolini, A., Mutulis, F., Wikberg, J.E., Schio¨th, H.B., 1998. Differential influence of a selective melanocortin MC4 receptor antagonist (HS014) on melanocortin-induced behavioral effects in rats. European Journal of Pharmacology 362 (2e3), 95e101. doi:10.1038/267261a0.
Vincent, J.D., 1994. Biology of pleasure. Presse Medicale 23 (40), 1871e1876. Vishnivetz, B., 1995. Educac¸ao do corpo para o ser. Editora Summus, Sao Paulo.
Walshe, F.M.R., 1923. On certain tonic or postural reflexes in hemiplegia with special reference to the so called «associated movements. Brain 46, 1e37.
Walusinski, O., 2006. Yawning: unsuspected avenue for a better understanding of arousal and interoception. Medical Hypotheses 67 (1), 6e14. doi:10.1016/j.mehy.2006.01.020.
Walusinski, O., Kurjak, A., Andonotopo, W., Azumendi, G., 2005. Fetal yawning assessed by 3D and 4D sonography. The Ultrasound Review of Obstetrics and Gynecology 5 (3), 210e217. doi:10.1080/14722240500284070.
Walusinski, O., Meenakshisundaram, R., Thirumalaikolundusubramanian, P., Diwakar, S., Dhanalakshmi, G., 2010. Yawning: Comparative Study of Knowledge and Beliefs, Popular and Medica. In the Mystery of Yawning in Physiology and Disease. Available at. http://www.baillement.com/recherche/beliefs_knowledge.pdf.
Wolff, J., 1986. The Law of Bone Remodelling. Springer-Verlag, Berlim.